Résumé
Un modèle théorique qui n’évolue pas avec de nouvelles informations issues de la recherche scientifique, en changeant les hypothèses dont il est né, devient une philosophie ; le scientifique devient un chef d’école. Même si, du clinicien au praticien de santé non médecin, elle est couramment pratiquée, la médecine ostéopathique crânienne manuelle est très controversée. Cet article, divisé en deux parties, passe en revue les hypothèses avec lesquelles le modèle crânien a été créé, mettant en évidence les innovations scientifiques et les nouvelles réflexions anatomo-physiologiques. Dans la première partie, nous passerons en revue la synthèse et le mouvement du liquide céphalo-rachidien (LCR), le mouvement du système nerveux central et périphérique ; nous mettrons en évidence les caractéristiques mécaniques des méninges. L’objectif de l’article est de souligner la nécessité de renouveler le modèle crânien existant.
Introduction et contexte
La médecine ostéopathique manipulative crânienne (OCMM) est née vers 1898, lorsqu’un étudiant de l’American School of Osteopathy a observé un crâne disjoint dans le musée du même institut et a remarqué que les os temporaux ressemblaient aux branchies du poisson [1]. L’étudiant en ostéopathie a vu que les contours osseux étaient construits pour rejoindre d’autres os en une articulation complexe. La similitude avec les branchies et la forme osseuse permettant les articulations du crâne a inspiré les principes fondamentaux de l’OCMM : le mécanisme respiratoire primaire (MRP) et le mouvement des os crâniens [1]. L’étudiant était William Garner Sutherland, futur DO. Le MRP est la théorie cherchant à expliquer le mouvement des os crâniens perçus à la palpation [2], Cette théorie repose sur cinq principes : la fluctuation du liquide céphalo-rachidien (LCR) ; la motilité inhérente du système nerveux central et de la moelle épinière ; la mobilité des membranes méningées (crâniennes et rachidiennes) ; mobilité articulaire des os du crâne ; le mouvement involontaire (passif) du sacrum entre les os iliaques [3]. Toujours selon cette théorie, le crâne possède un rythme ou respiration crânienne ; ce rythme a une oscillation palpable d’environ 12 cycles par minute [3]. Une autre pierre angulaire du MRP est le mouvement de l’articulation entre l’os occipital et l’os sphénoïde (synchondrose sphéno-basilaire ou SSB), un point d’appui important pour identifier les dysfonctionnements structurels de la forme et de la fonction du crâne [3], selon l’OCMM, les os de la ligne médiane comprennent l’os sphénoïde, l’occiput, l’ethmoïde et le vomer. Pendant le MRP du crâne, ceux-ci subiraient une flexion et une extension et seraient responsables du mouvement de rotation externe et interne des autres os crâniens respectivement appariés [3]. Lors de la flexion de la SSB, les méninges tirent l’os sacré vers le haut, entraînant une extension anatomique ou une contre-nutation de la base sacrée ; l’inverse se produit lors d’une extension de la SSB [3].
L’ostéopathe pose ses mains sur le crâne et évalue les éventuels dysfonctionnements crâniens [2]. Un dysfonctionnement crânien fait référence à un mouvement perçu étranger à ce qui devrait être normal (flexion-extension ou rotation interne-externe) [1]. Classiquement, avec la palpation et suivant la théorie du MRP, diverses dysfonctions crâniennes spécifiques peuvent être identifiées au niveau de la SSB : cisaillement, déformations latérales, torsions, rotations en flexion latérale et compressions, entre autres [3]. D’autres anomalies crâniennes qui peuvent être palpées sont attribuables à un dysfonctionnement dural, comme les strains latéraux (droit ou gauche) et supérieurs ou inférieurs ; les lésions supplémentaires comprennent des strains intra-osseux et des mouvements anormaux des os du crâne individuels (29 os chez les adultes) [3-4]. Suite à de nombreuses publications de la Fondation de la recherche ostéopathique et de l’approbation clinique (FORCE) sur le tissu fascial, est né un groupe d’étude pour comprendre, améliorer et éventuellement créer un nouveau modèle d’OCMM [5-6]. Ce groupe, Cranial Research And New International Osteopathy : CRANIO, implique des ostéopathes et des chercheurs de différentes nationalités. Le travail que nous proposons est divisé en deux articles. La première partie a pour objectif de passer en revue et de comparer les connaissances scientifiques les plus récentes par rapport à la base théorique sur laquelle repose le MRP chez l’adulte, en essayant de mettre en évidence la nécessité d’un nouveau modèle d’OCMM. La seconde partie complétera la revue et tentera de proposer une nouvelle manière de concevoir l’OCMM, toujours à travers la littérature scientifique actuelle.
Production du liquide céphalo-rachidien (LCR)
Selon l’OCMM, le mouvement crânien commence par la synthèse et la distribution du LCR [7-8]. La présence de fluide entourant le cortex est connue depuis l’époque d’Imhotep, un médecin égyptien d’il y a 4000 ans, d’Hérophile (280 avant JC) et d’Hippocrate (370 avant JC), ainsi que de Galien de Pergame (200/216 après JC) [9]. Hippocrate fut le premier à décrire ce fluide comme de l’eau qui entoure le cerveau, jusqu’en 1764, année où le médecin italien Cotugno décrivit la présence d’eau mais en lui donnant un nom spécifique : cotunnii LCR [10-11]. La terminologie utilisée aujourd’hui, à savoir liquide céphalo-rachidien, n’apparaît qu’en 1842 dans les écrits de Magendie [11]. Le LCR a deux origines ontogéniques, selon le stade de développement de l’embryon. La plaque neurale, qui dérive d’une partie de l’ectoderme dorsal, est en contact avec le liquide amniotique (LA) ; la plaque neurale se plie pour former le tube neural avec un vecteur longitudinal [12]. Ce mouvement emprisonne une partie du LA à l’intérieur du tube neural. Le tube neural primitif est en contact avec le LA, grâce à l’ouverture des neuropores, qui ne sont pas encore fermés ; cela signifie que son développement est affecté par l’information fluidique du LA [12]. Le tube neural subit d’autres transformations. Dans sa partie postérieure, il formera la future moelle épinière tandis que, dans la zone antérieure, il se dilatera pour former le système nerveux central [12]. Les placodes neurogènes resteront en contact avec le LA tandis que le système cérébral naissant entrera en contact avec le fluide qui deviendra le LCR en passant par le LA [13]. Aux premiers stades de la formation du cerveau, ce dernier est un espace vide rempli de fluides et entouré d’un neuroépithélium primitif monocouche. Lorsque les neuropores antérieurs se ferment définitivement, la zone cérébrale est indépendante dans la production de LCR et ne reflète plus les changements métaboliques de l’environnement fluide externe [12]. La couche précurseure du neuroépithélium se développe davantage, suite à une histogenèse spécifique. Au cours de l’embryogenèse, le LCR ne circule pas mais facilite plutôt le gonflement et l’élargissement des espaces qui seront comblés par les structures du système nerveux ; les capillaires, jouent alors un rôle fondamental dans la sécrétion et la résorption du LCR [14]. Pendant cette période de formation, le système mésodermique produit le LCR (vaisseaux sanguins). À la fin de la période embryonnaire, c’est-à-dire avec l’apparition des esquisses des plexus choroïdes, le LCR est produit par des structures ectodermiques [15]. Les plexus choroïdes deviennent un important centre de production de LCR. La neurogenèse commence lorsque les plexus commencent à fonctionner [15]. Les molécules présentes dans le LCR du fœtus et de l’adulte sont différentes et n’ont pas les memes fonctions [15]. Jusqu’à l’apparition des plexus (41 jours de gestation) dans le quatrième ventricule, le LCR ne circule pas [15]. Une autre différence du LCR fœtal est que ce dernier agit pour créer des forces expansives, tandis que celui de l’adulte maintient les forces fluidiques en équilibre [15]. L’un des points clés du rythme crânien est la production de LCR à partir des plexus choroïdes (à partir du plasma sanguin), puis sa libération dans les ventricules. En réalité, il existe d’autres sites de synthèse, comme les organes circumventriculaires, la couche neuroépithéliale qui recouvre les ventricules, quoique dans une moindre mesure [9]. La production quotidienne moyenne de LCR est de 400 à 600 ml, avec un volume constant d’environ 150 ml et une pression moyenne chez les adultes d’environ 10 à 15 mmHg ; la production est influencée par divers facteurs, dont l’innervation des plexus choroïdes [14]. Le système sympathique a tendance à réduire la sécrétion de LCR tandis que le stimulus parasympathique en stimule la synthèse [14]. Il a été démontré que certains neuropeptides (avec des récepteurs spécifiques sur les plexus) jouent un rôle dans la production, comme la sérotonine, la dopamine, la mélatonine, ainsi que le peptide natriurétique auriculaire (ANP) et l’arginine vasopressine (AVP) [14]. L’ANP est sécrétée par les myocytes dans les oreillettes du myocarde, en particulier, dans le but principal de réduire la pression artérielle, tandis qu’au niveau des plexus, en activant le canal de l’aquaporine-1 (AOP1), il est capable de réduire la synthèse du LCR [16]. Les maladies cardiaques, telles que l’insuffisance cardiaque chronique (ICC), pourraient affecter la production de LCR. L’AVP est synthétisée dans certaines zones cérébrales et par l’épithélium des plexus choroïdiens, en particulier lorsqu’il y a une modification de la quantité d’eau dans le corps, impliquant les canaux Na (+) voltage-dépendants et des récepteurs spécifiques tels que le récepteur AVP VI ; son action sur les plexus est de stimuler la réduction de la production de fluide [17]. Étant donné que l’AVP est également connue sous le nom de molécule « sociale », les sautes d’humeur pourraient induire des changements dans la production de liquide.
Selon certaines recherches, le parenchyme cérébral est une autre source de production, notamment en raison de l’apport en eau [18]. Une information scientifique importante sur le comportement du LCR est la variation de la zone de production, en fonction de la posture de la personne. Il a été montré que si la personne est en décubitus dorsal, la synthèse provient notamment des cellules épendymaires de la moelle épinière tandis que si la personne est debout, la synthèse du LCR se fait principalement à partir des plexus choroïdes [19-20]. La synthèse du LCR ne se produit pas seulement dans les ventricules, mais également dans d’autres zones anatomiques ; la synthèse dépend de la posture et des différentes molécules et systèmes nerveux qui influencent le comportement des plexus choroïdes. Cela dépend de la présence de lumière et d’obscurité (rythme circadien). La mélatonine, en plus de la production par les pinéalocytes (qui sont en contact avec le LCR), est produite par les tanycytes, cellules épendymaires spécialisées que l’on trouve dans le troisième ventricule ; la mélatonine synthétisée la nuit semble avoir la capacité d’augmenter la synthèse du LCR, de 12 mL par heure pendant la journée, jusqu’à un maximum de 42 mL par heure la nuit [19, 21-22]. Le LCR est également synthétisé par le parenchyme cérébral et la pie-mère de la surface cérébrale [23]. La synthèse du LCR n’est pas un processus passif mais répond à de multiples variables de contrôle homéostatique, par rapport à l’environnement dans lequel il circule et par rapport aux variations de l’environnement.
Mouvement du liquide céphalo-rachidien
Un autre concept qui crée une dichotomie avec le modèle ostéopathique du MRP concerne le mouvement du liquide cérébral et rachidien. La circulation du LCR n’est pas unidirectionnelle et son écoulement (direction, force, volume, vitesse) n’est pas uniforme. Le débit ne correspond pas à la quantité de LCR produit et peut être bidirectionnel et oscillatoire [24]. On pourrait comparer son flux aussi aléatoire et hétérogène à un tableau de Pollock. Dans le système nerveux central et rachidien, bien que nous en sachions peu et qu’il existe des études sur les animaux et peu sur l’homme, le déplacement du LCR est également stimulé par certaines structures appelées fibres de Reissner (RF). Les RF consistent en l’agrégation d’une glycoprotéine (scospondine) et, bien qu’elles jouent un rôle important au cours de l’embryogenèse, certaines preuves mettent en évidence le rôle dans la facilitation de l’écoulement du fluide ; chez l’adulte, elles auraient probablement un rôle de capteur mécanotransductif au passage du LCR [25-26]. L’absorption du LCR par les villosités arachnoïdiennes et son écoulement vers le système veineux, comme généralement enseigné dans le cadre de l’ostéopathie crânienne, n’est pas si vrai [23]. Dans des conditions physiologiques, la voie de circulation la plus importante pour que le LCR sorte du crâne est le système lymphatique nasal [23]. L’espace périvasculaire qui entoure les veines et les artères au niveau sous-arachnoïdien, et pénètre et entoure le parenchyme cérébral, est appelé espace de Virchow-Robin [10]. Les mécanismes qui permettent le mouvement du LCR dans cet espace sont liés aux pulsations artérielles et à un mécanisme appelé convection ; ce dernier est le mouvement des fluides à travers la relation entre la chaleur du fluide lui-même et son gradient de masse (Figure 1) [27].
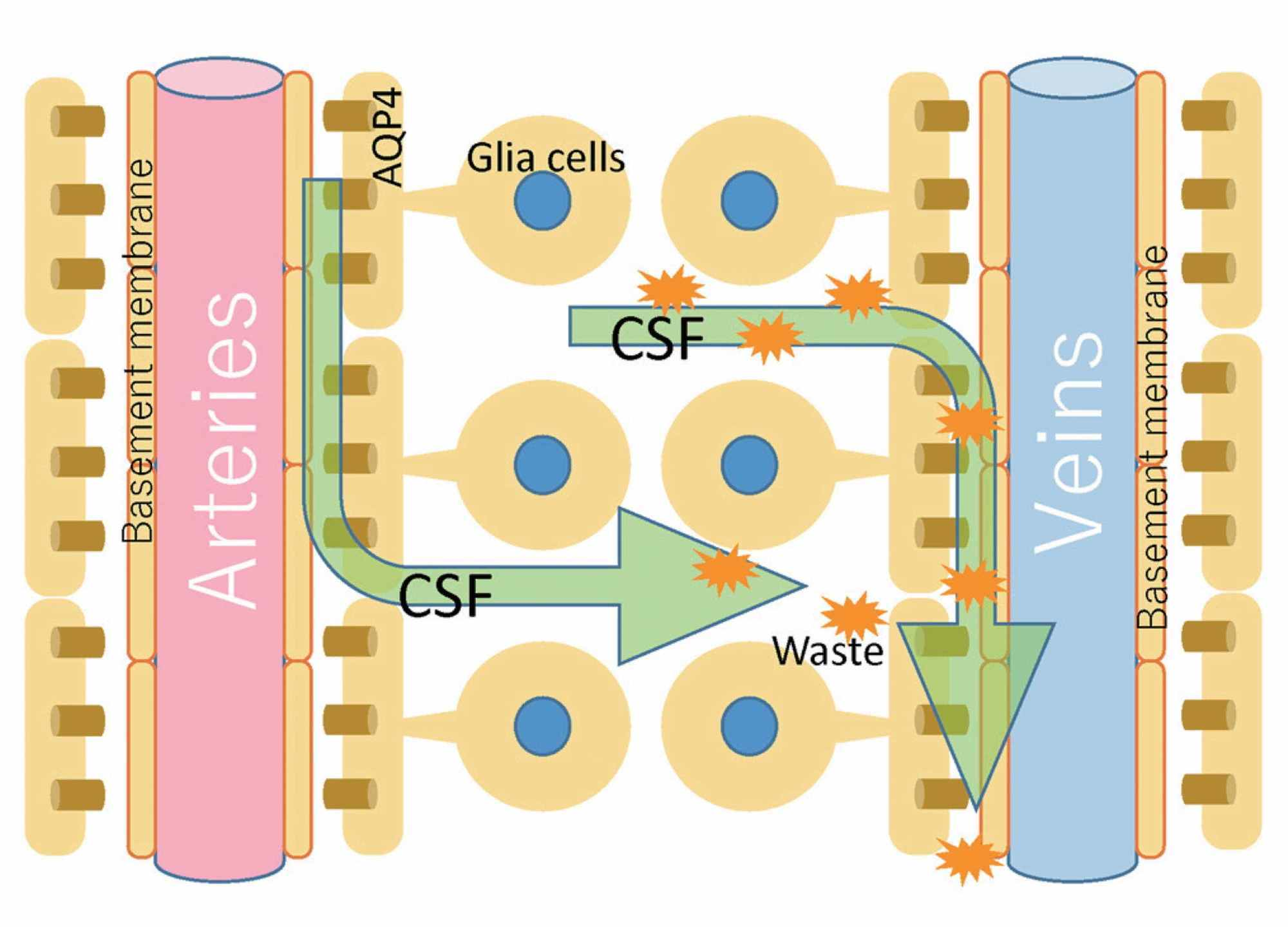
Figure 1 : La figure illustre schématiquement ce que pourrait être la distribution du LCR à travers les espaces périvasculaires, en mélange avec les fluides interstitiels.LCR, liquide céphalo-rachidienImage reproduite avec la permission de Toshiaki Taoka, MD, PhD et collègues, Département de visualisation biomédicale innovante, Nagoya, Japon.
De plus, le LCR peut pénétrer dans l’espace périvasculaire grâce aux pores des vaisseaux (notamment au niveau de l’espace sous-arachnoïdien), appelés stomates (Figure 2) [21].
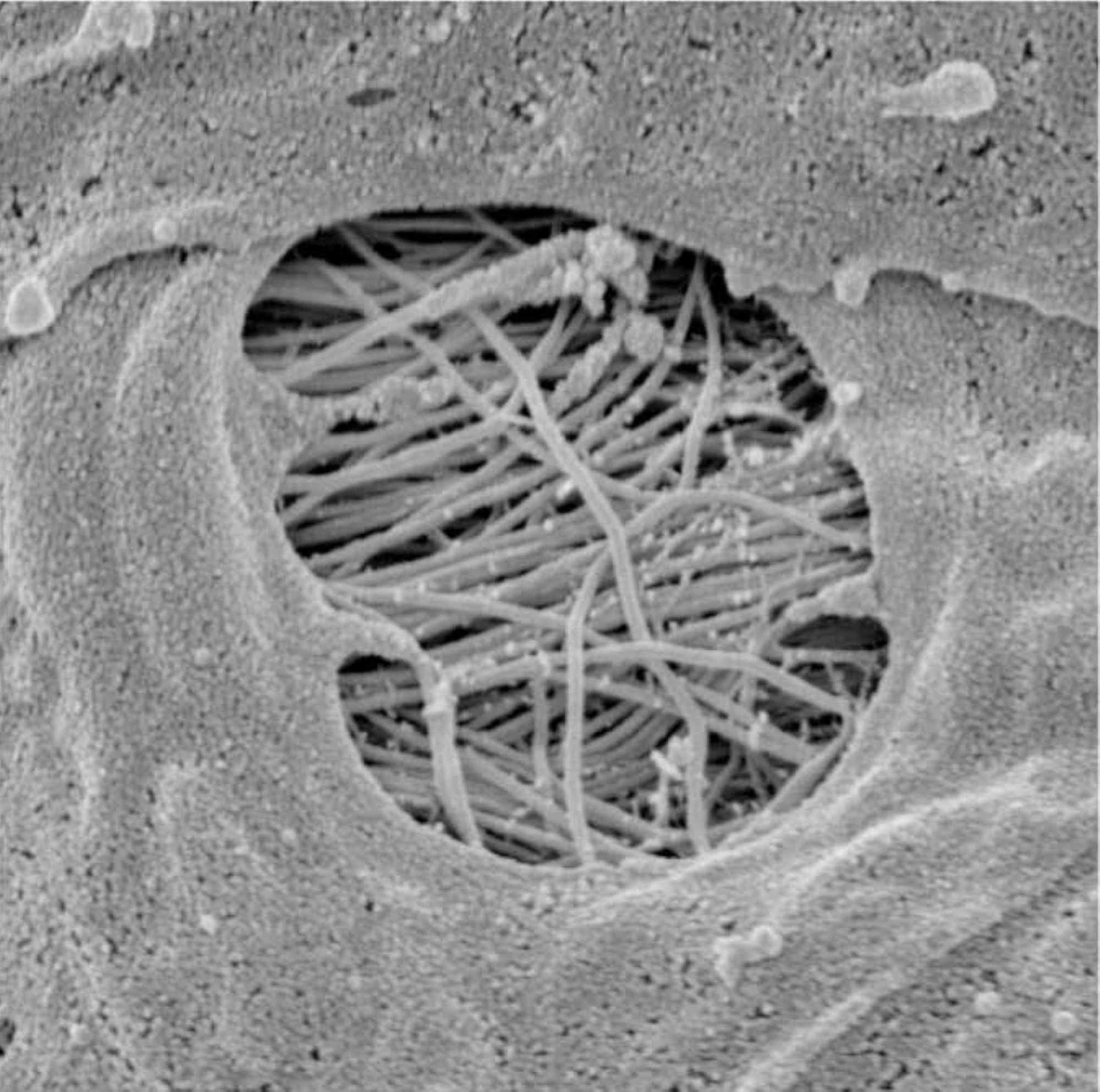
Figure 2 : L’image montre un pore ou des stomates d’un vaisseau dans l’espace sous-arachnoïdien dans lequel le LCR peut pénétrer dans l’espace périvasculaire.LCR, liquide céphalo-rachidienImage reproduite avec la permission du professeur Joan Abbott, Faculté des sciences de la vie et de la médecine, Institut des sciences pharmaceutiques, King’s College, Londres, Royaume-Uni.
Le LCR, ayant pénétré dans l’espace artériel périvasculaire, pénètre dans l’interstitium cérébral, grâce à la régulation des protéines aquaporines (aquaporine 4 – A0P4) des canaux d’eau (le LCR est à 99 % d’eau) ; dans ces canaux se trouvent les parties terminales des astrocytes ou cellules gliales, qui constituent la paroi externe de l’espace périvasculaire [23]. Dans ce processus, le LCR se mélange à l’espace interstitiel, échangeant des solutés (avec des mécanismes de diffusion), des molécules et nettoyant les différents résidus métaboliques [23]. Une fois le liquide interstitiel nettoyé, le LCR se déplace vers l’espace veineux périvasculaire [23]. Le mécanisme d’échange entre les fluides interstitiels et le LCR au niveau des espaces périvasculaires est défini comme le système glymphatique (cellules gliales et lymphatique) [21]. On sait qu’il existe des vaisseaux lymphatiques méningés, mais il est probable qu’ils n’ont pas d’échanges directs avec le LCR, sinon hors du système nerveux [28]. Les voies suivies par le LCR à la sortie du crâne sont variées. De l’espace périvasculaire veineux, le LCR atteint le bulbe olfactif, pour entrer dans l’espace périneural du nerf olfactif (1596-2096 de tout le LCR), jusqu’à la muqueuse nasale, où le LCR sera drainé par les vaisseaux lymphatiques de la muqueuse ; à partir de là, le liquide pénètre dans les voies lymphatiques sous-buccales et mandibulaires, jusqu’aux ganglions lymphatiques cervicaux et rachidiens [28]. À la base du crâne, de nombreux vaisseaux lymphatiques pénètrent dans le crâne en suivant les voies de sortie des nerfs crâniens et spinaux [11]. Le LCR a probablement la possibilité de pénétrer dans ces vaisseaux périphériques et, à nouveau, de pénétrer dans les ganglions lymphatiques du tractus cervical, en suivant également les voies vertébrales [11]. Le LCR est également absorbé par l’oreille interne, la carotide interne, l’espace périneural des nerfs crâniens, les nerfs spinaux et intercostaux et le système veineux crânien [11, 21]. Le LCR qui suit l’espace périneural vers le tissu musculaire pourrait probablement entrer en contact avec les fluides interstitiels musculaires.
Pulsation du liquide céphalo-rachidien
Les principales forces documentées pour déterminer le mouvement du LCR sont le rythme cardiaque et la respiration diaphragmatique ; le cœur et le muscle diaphragme créent les pulsations du LCR. Lors d’une inspiration, le LCR est poussé crânialement (de l’espace rachidien périvasculaire et périneural vers le crâne), tandis que lors d’une expiration, le mouvement se fait caudalement [19]. En décubitus dorsal, la pression intracrânienne du LCR est la plus faible (4,6 mmHg) par rapport aux autres postures et malgré les forces centripète et centrifuge [19]. Il est probable que la respiration agit principalement sur la voie veineuse périvasculaire mais des études plus approfondies sur l’homme font défaut [29]. La systole et la diastole sont capables de déplacer le LCR, mais de manière moins importante que le muscle diaphragme. On suppose que le rythme cardiaque influencerait le plus le mouvement dans l’espace périvasculaire artériel [29]. Pendant la systole, le LCR est poussé caudalement, tandis que, avec la diastole, le LCR est poussé crânialement. Il faut se rappeler que le mouvement du LCR, grâce à la respiration et aux battements rythmiques du cœur, n’est jamais complet dans tout le corps (il ne fait pas un tour complet) ; le mouvement lui-même stimule les ondes, les rythmes qui permettent au LCR de se déplacer dans le système corporel [24]. L’unidirectionnalité du LCR n’existe pas alors que les pulsations du LCR se retrouvent dans tout le corps [24]. Une structure qui pourrait aider au déplacement bidirectionnel du LCR entre la base du crâne et la moelle épinière cervicale est une lame de tissu arachnoïdien de morphologie rhomboïde, entre la moelle allongée et les racines cervicales (C4) [30]. Elle est appelée « valva cerebri » pour son rôle hydrodynamique pour le mouvement du LCR ; des études complémentaires sont nécessaires [30]..
Motilité inhérente du système nerveux central et de la moelle épinière
Selon le concept d’OCMM, le système nerveux central et périphérique aurait un mouvement intrinsèque ou inhérent, capable de déplacer le LCR et les méninges crâniennes [1-3]. Ces mouvements détermineraient le rythme du MRP et, par conséquent, le rythme ressenti par l’évaluation palpatoire de l’ostéopathe (10-14 cycles) [8]. De nombreux éléments et forces sont capables d’influencer la motilité du système nerveux. La matrice extracellulaire qui imprègne le cerveau et la moelle épinière (et tout le corps) possède des propriétés piézoélectriques. Un stimulus électrique voyageant à travers la matrice provoquera une vibration, tandis que le passage d’informations mécaniques-métaboliques provoquera la création d’électricité [31]. Ces changements de comportement de la matrice extracellulaire ou les fluctuations d’énergie affecteront l’état de la matière de la même matrice (gélification et solution), où l’eau présente se comporte comme un cristal liquide [31]. Les ions « électrifiés » de la matrice influençant la vitesse des fluides (électro-osmose) ; la vitesse ne dépend pas des vannes ou des pompes mais uniquement de la charge électrique des ions [32]. Les réactions biochimiques entre les neurones et les cellules qui composent le complexe nerveux peuvent créer des ondes chimiques sous l’égide de la loi de la thermodynamique hors équilibre, ce qui impliquerait notamment le centrosome (MicroTubule Organizing Center – MTOC) [33]. Le centrosome réorganise la forme de la cellule, modifie la tension de la cellule elle-même et fait varier la tension mécano-électrique de la matrice extracellulaire, repropose le mécanisme de gélification (gel ou phase liquide interconnectée) ou de solution (sol ou suspension colloïdale plus solide), influençant la vitesse des fluides [33]. Ces mécanismes ne sont pas homogènes dans le système nerveux central ou périphérique et ne sont donc pas capables d’influencer la mobilité du complexe nerveux dans son intégralité. Certains neurones peuvent créer des nanotubes pour le transport des mitochondries et d’autres éléments structurels ou molécules vers un ou plusieurs neurones, faisant varier la tension électromécanique des zones cérébrales, modifiant l’état de vitesse des fluides cérébraux [34]. Cela étant, il n’y a pas d’homogénéité dans tout le système nerveux, et il faut souligner que l’existence des nanotubes dure de quelques minutes à quelques heures [34]. Selon le principe de l’intrication quantique, toutes les cellules sont en communication les unes avec les autres, comme une sorte de câblage ou de transmission éphaptique, mais le mouvement produit (oscillation, vibration, déformation morphologique) dépend toujours de l’adaptation d’une région cérébrale, par rapport à un autre [34-35]. Ces mécanismes ne sont pas capables de déplacer les systèmes nerveux central et périphérique à l’unisson. Une autre force qui agit sur le système nerveux est la vasomotricité, contrôlée par les systèmes nerveux sympathique et parasympathique et par la même structure des cellules contractiles qui forment le vaisseau, ces dernières étant influencées par les événements métaboliques. Selon une vision scientifique ostéopathique, la modification du tonus des vaisseaux sanguins (et lymphatiques) serait à la base du rythme crânien [8]. Les oscillations sont liées à la pression artérielle et aux différents neurofluides, qui sont historiquement mais de manière incohérente liés à des ondes mesurables. On connaît les ondes Traube-Hering (0,1 Hz) liées à la pression artérielle mais, qui, en réalité, ne reflètent pas la vitalité du rythme crânien que ressent l’ostéopathe avec l’évaluation manuelle, car 0,1 Hz correspond à des oscillations de pathologie (perte d’autorégulation autonome) [36]. Une autre oscillation plus ample (0,005-2 Hz) est influencée par le LCR dans l’espace sous-arachnoïdien, mais on sait que le déplacement du LCR est hétérogène et ne peut pas représenter un rythme homogène tel que celui ressenti par la palpation de l’ostéopathe [36]. De plus, cette oscillation n’est pas égale entre les deux hémisphères et peut être influencée par l’état émotionnel du patient [36]. Une autre oscillation connue et mesurable est celle de Mayer (0,1 Hz), mais sa nature reste méconnue et douteuse ; ces oscillations sont probablement liées à l’activité nerveuse autonome et aux déplacements des fluides interstitiels [37]. Des oscillations de Mayer peuvent également être enregistrées dans la moelle épinière, probablement avec les mêmes raisons que celles émises pour le système nerveux central [38]. Un mécanisme capable de déplacer de manière homogène le système nerveux central et le système nerveux rachidien est celui lié au cœur et au muscle diaphragme. Nous savons que le système nerveux central et la moelle épinière se déplacent de 2 à 3 ml, stimulés par le diaphragme respiratoire et le rythme cardiaque. Pendant la systole, le système nerveux est tracté caudalement et médialement ; l’inverse se produit pendant la diastole [8]. Lors d’une inspiration, le système nerveux est poussé vers le crâne, tandis que lors d’une expiration, le mouvement prédominant est caudal [8]. Les systèmes cardiovasculaire et respiratoire fonctionnent en parfait équilibre chez un sujet sain et sont contrôlés par les mêmes systèmes neurologiques (et métaboliques) : système vagal, système sympathique, système phrénique [39]. La fréquence cardiaque varie avec le rythme de la respiration, tout comme la fréquence respiratoire est affectée par le comportement du myocarde [39], Selon les informations scientifiques les plus récentes, le mécanisme de la respiration primaire (mouvement), ne peut pas provenir directement de la masse cérébrale et de la moelle épinière, mais il s’agit d’une réponse (mouvement) déterminée par la respiration / au rythme cardiaque. Dans le domaine de l’OCMM, il serait plus juste de parler du mécanisme respiratoire secondaire (MRS). Il est probable que lorsque l’ostéopathe pose ses mains sur le crâne pour une écoute palpatoire, il est principalement conscient de la santé du système cardiorespiratoire.
Mobilité des membranes méningées crâniennes et spinales
Les méninges crâniennes sont la faux du cerveau, la tente du cervelet, la faux du cervelet et la tente de l’hypophyse [7-8]. La dure-mère du crâne, qui recouvre la couche arachnoïdienne et a une épaisseur moyenne d’un millimètre, a des propriétés viscoélastiques anisotropes, avec une capacité à résister à des forces mécaniques d’environ 9-10 Pa (MPa qui dans ce cas, équivaut à 1 kilo/millimètre2) [40]. La dure-mère est définie comme pachyméninge, tandis que l’arachnoïde et la couche piale relèvent de la classification de leptoméninge. Les méninges touchant la zone du cerveau antérieur caudal et du mésencéphale ont une origine ectodermique, tandis que les méninges restantes proviennent du mésoderme (comme les vaisseaux des méninges) [41]. Plus précisément, la dure-mère dérive du mésoderme, tandis que la pie-mère et l’arachnoïde dérivent du mésoderme et de l’ectoderme [41]. La couche externe de la dure-mère ou couche périostée durale est en contact direct avec le périoste du crâne, avec lequel elle échange des vaisseaux artériels [41]. La couche arachnoïdienne a une épaisseur d’environ 200 µm ; la pie-mère est une couche unique de cellules ou de membrane basale, avec une matrice extracellulaire spécialisée recouvrant le parenchyme cérébral et les vaisseaux qui pénètrent dans le cerveau [41]. Si l’arachnoïde agit comme un amortisseur et comme un élément dispersif de tension mécanique, la pie-mère répond également aux contraintes mécaniques et aux stimuli nociceptifs [42]. Les méninges sont innervées par le système autonome (systèmes vagal et sympathique), par les nerfs crâniens (IV, V, VII, IX, XII) et par les quatre premières racines cervicales [7-8]. Les méninges sont importantes pour l’écoulement veineux, de la dure-mère avec les sinus veineux, jusqu’à la pie-mère avec les veines pontantes (ces dernières reliées aux sinus duraux), en passant par la couche arachnoïdienne (Figure 3) [43].
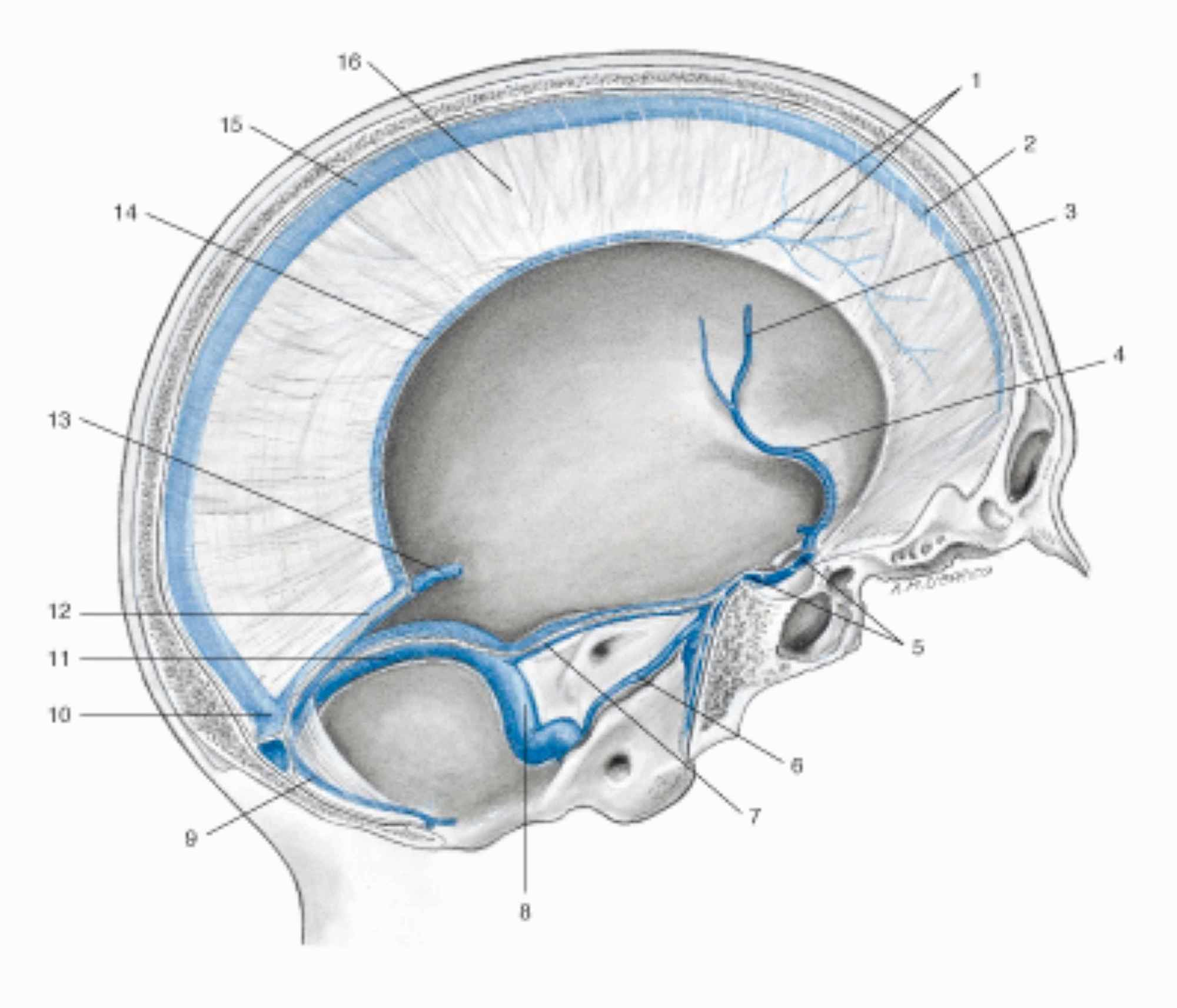
Figure 3 : Représentation des sinus veineux de la dure-mère dans une coupe sagittale du crâne. 1. Veines de la dure-mère ; 2. Sinus sagittal supérieur ; 3. Veine cérébrale moyenne gauche ; 4. sinus sphénopariétal gauche ; 5. Sinus intercaverneux ; 6. Sinus pétreux inférieur gauche ; 7. Sinus pétreux supérieur gauche ; 8. Sinus sigmoïde gauche ; 9. Sinus occipital ; 10. Confluence des sinus ; 11. Sinus transverse gauche ; 12. sinus droit ; 13. Grande veine cérébrale (de Galien) ; 14. Sinus sagittal inférieur ; 15. Sinus sagittal supérieur ; 16. Faux du cerveau.Image reproduite avec autorisation, d’après Anastasi G, et al., Anatomia dell’uomo, quatrième édition [Human Anatomy], 2010, Milan : Edi-Ermes, Volume , Figure 4.145, p. 432.
Le système méningé est riche en vaisseaux lymphatiques qui, du point de vue du développement, ne se forment qu’après la naissance ; ils suivent le trajet des sinus veineux et le trajet de certaines artères, comme l’artère méningée moyenne [44]. Les méninges transmettent des forces mécaniques de l’extérieur vers l’intérieur (traumatisme), et de l’intérieur vers l’extérieur ; de plus, ce sont des structures qui reflètent les variables mécano-métaboliques de l’environnement dans lequel elles résident, modifiant leur capacité à gérer les tensions et faisant varier leur structure intrinsèque dans le temps [8]. Avec l’âge, la dure-mère a tendance à s’ossifier, notamment au niveau des sinus de la dure-mère, ce qui affectera la transmission des efforts mécaniques [8]. Le repli de la dure-mère pour former les sinus veineux suit la morphologie du cerveau, qui n’est jamais parfaitement la même dans les deux hémisphères ; il s’ensuit que les sinus peuvent être d’orientation spécifique (droite ou gauche), avec une rigidité tissulaire anisotrope [8]. Nous ne disposons pas de données détaillées sur le comportement méningé et le mode de transmission des tensions détectées, mis à part le fait qu’elles agissent comme des amortisseurs et qu’elles peuvent élargir ou réduire la propagation des tensions (pressions). En regardant les études sur le comportement des tissus entourant le cerveau en présence d’un traumatisme, il est clair que si la force est externe et implique le crâne, la tension produite est amplifiée ; de la peau au parenchyme, les différentes couches augmentent la force de pression vers le cerveau [45]. En se référant aux données actuelles, on pourrait supposer que si une tension (pression ou traction) vient du cerveau vers la peau, les différentes couches devraient amortir et ralentir la vitesse de propagation des forces de tension produites [45]. En effet, si un vecteur de contrainte traverse des couches de rigidité (osseuse) importante, la vitesse des ondes de propagation augmente (de l’extérieur vers l’intérieur) ; à l’inverse, si les forces sont produites de l’intérieur, par exemple lorsque le cerveau et la moelle épinière bougent sous l’influence de la respiration et du rythme cardiaque, la vitesse de propagation des pressions créées est plus lente, car les tissus internes ont moins de rigidité [46]. Il est probable que le rythme crânien ressenti par la palpation du clinicien dérive du mouvement du cerveau et de la moelle épinière qui créerait des ondes de tension vers l’extérieur plus lentes, par rapport au rythme réel de la respiration et du rythme cardiaque, grâce à la propriété viscoélastique des méninges qui amortit la vitesse de transmission de tension. La dure-mère spinale, d’origine embryologique mésodermique, enveloppe la moelle épinière dans son intégralité ; lorsque les neurofluides traversent la zone protégée par la dure-mère, celle-ci s’étire et appuie contre la zone vertébrale [47]. La dure-mère crânienne se poursuit dans la moelle épinière, du foramen magnum à la région périostée vertébrale ; au niveau de la deuxième vertèbre sacrée (S2), elle se poursuit avec une structure filiforme, appelée filum terminale, qui relie le sac dural au sacrum (périoste) [47]. Dans l’espace sous-dural existent des structures qui stabilisent la relation de la dure-mère avec la moelle épinière (septum posticum fibreux) ; la dure-mère rachidienne est innervée notamment au niveau ventral et moins au niveau dorsal, par les nerfs sinuvertébraux et par des branches du système sympathique [47], La dure-mère vertébrale est construite pour mieux résister aux tensions longitudinales et dans une moindre mesure aux tensions latérales. La couche durale interne est en continuité avec la couche arachnoïdienne ; l’espace sous-arachnoïdien est symétrique sur les côtés, mais présente des asymétries entre la zone ventrale et dorsale [47], Le sac dural peut se déplacer dans le sens longitudinal de 2-4 mm, une mesure qui coïncide avec le mouvement du système nerveux [47], Le système lymphatique rachidien se trouve dans l’espace péridural, avec le tissu adipeux [48], Malgré sa faible épaisseur (de 0,089 à 1,40 MPa), la couche spinale piale a une grande capacité à résister aux contraintes mécaniques longitudinales et à leurs distributions [ 49-50].
Conclusions
L’OCMM s’appuie sur la palpation du crâne pour évaluer et éventuellement résoudre les changements de mouvement, de forme et d’orientation des os crâniens. La revue de l’article, divisée en deux parties, discute des données récentes sur la synthèse et le mouvement du LCR, met en évidence le mouvement du système nerveux central et les caractéristiques mécaniques des méninges crâniennes et spinales.
De la première partie de l’article il est possible de déduire de nouvelles réflexions, comme celle de nommer le MRP avec le nom de MRS, et la possibilité que le mouvement perçu par les mains de l’ostéopathe placées sur le crâne, dérive du mouvement du cerveau (grâce au cœur et au diaphragme respiratoire), filtré par l’amortissement des méninges crâniennes.
Bordoni B, Walkowski S, Ducoux B, et al.
distribué sous les termes de la licence d’attribution Creative Commons CC-BY 4.0
10.7759/cureus.10410
Traduction par Pierre Tricot
Information additionnelle
Divulgations
Conflits d’intérêts : Conformément au formulaire de divulgation uniforme de l’ICMJE, tous les auteurs déclarent ce qui suit : Informations sur le paiement / les services : Tous les auteurs ont déclaré qu’aucune organisation n’avait reçu de soutien financier pour le travail soumis. Relations financières : tous les auteurs ont déclaré n’avoir aucune relation financière à l’heure actuelle ou au cours des trois années précédentes avec des organisations qui pourraient avoir un intérêt dans le travail soumis. Autres relations : Tous les auteurs ont déclaré qu’il n’y avait aucune autre relation ou activité qui pourrait sembler avoir influencé le travail soumis.







